Arbeitet eine Zelle unter anaeroben Bedingungen, produziert sie Energie, indem sie Glukose in Laktat umwandelt und über den Cori-Zyklus diese entsorgt; wenn Sauerstoff zur Verfügung steht (also im Ruhezustand), werden mehr als 90 % der Glukose aerob verbraucht und nur die restlichen 10 % anaerob. Wenn mehr ATP benötigt wird, als der aerobe Weg liefern kann (z beim Atmen usw.): Dieser Stoffwechsel muss beschleunigt werden, indem Laktat (das aus der Glykolyse gewonnen wird) durch Glukoneogenese in Glukose umgewandelt wird.
In den Mitochondrien entwickelt sich der aerobe Stoffwechsel.
Das erste Enzym, das im aeroben Stoffwechsel vorkommt, ist das Pyruvatdehydrogenase; Genauer gesagt ist die Pyruvatdehydrogenase eher ein Enzymkomplex als ein Enzym, da sie ein Aggregat von 48-60 Proteineinheiten mit drei nacheinander wirkenden katalytischen Zentren ist.
Pyruvat-Dehydrogenase katalysiert folgende Reaktion (Redox):
Pyruvat + NAD + + CoA-SH → Acetyl-CoA + NADH + H + + CO2
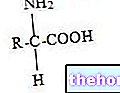
CoA-SH ist Coenzym A: Es ist ein Derivat der Pantothensäure, Acetyl-Coenzym A ist ein Thioester. Dies ist ein Redox-Prozess, da der erste Kohlenstoff von Pyruvat von Oxidationszahl drei zu Oxidationszahl vier (es oxidiert) und der zweite Kohlenstoff von Pyruvat von Oxidationszahl zwei zu Oxidationszahl drei (es oxidiert) geht. Dann wird das Pyruvat oxidiert (es verliert insgesamt zwei Elektronen) und das NAD wird reduziert.
Wie bereits erwähnt, weist Pyruvat-Dehydrogenase drei Arten von enzymatischer Aktivität auf, die jeweils durch ihren eigenen katalytischen Cofaktor unterstützt werden:
- Thiaminpyrophosphat (es ist ein Derivat von Vitamin B1); es ist in deprotonierter Form aktiv: Es entsteht ein Carbanion.
- Lipoamid (es ist ein Derivat der Liponsäure); es enthält eine sehr reaktive Disulfidbrücke.
- Flavinadenindinukleotid (es ist ein Derivat von Vitamin B2); es ist ein Nukleotid mit Redoxeigenschaften: Sein Redoxzentrum besteht aus dem Flavin.
In eukaryotischen Zellen findet der aerobe Metabolismus in spezialisierten Organellen der Zelle statt, die die Mitochondrien sind; bei Bakterien findet der Stoffwechsel von Glukose und anderen Spezies in der Zelle statt, aber es gibt keine spezialisierten Organellen.
Wenn Pyruvat in ein Mitochondrium eindringt, wird es der "Wirkung der Pyruvat-Carboxylase" ausgesetzt, wenn eine Glukoneogenese durchgeführt werden muss (um das Ausgangsmaterial zu rekonstruieren), oder es kann der Pyruvat-Dehydrogenase ausgesetzt werden, wenn es zur Energiegewinnung erforderlich ist: Acetyl-Coenzym A, das durch aeroben Stoffwechsel gebildet wird, stimuliert die Wirkung der Pyruvat-Carboxylase, fördert daher die Gluokoneogenese und reduziert die Wirkung der Pyruvat-Dehydrogenase.
Sehen wir uns nun an, wie Pyruvat-Dehydrogenase funktioniert; Zunächst erfolgt eine Decarboxylierung von Pyruvat durch die Wirkung von Thiaminpyrophosphat.
Eine saure Umgebung kann den aeroben Stoffwechsel hemmen, da die anionische Form von Thiaminpyrophosphat aktiv ist, die bei saurem pH protoniert würde und keine Decarboxylierung stattfinden würde.
Eine Decarboxylierung ist eine schwierige Reaktion, da eine Kohlenstoff-Kohlenstoff-Bindung aufgebrochen werden muss; in diesem Fall wird die Reaktion thermodynamisch dadurch begünstigt, dass das Reaktionszwischenprodukt (Hydroxyethyl-Thiamin-Pyrophosphat) Resonanz erzeugt (die p-Elektronen des Moleküls sind delokalisiert): das Hydroxyethyl-Thiamin-Pyrophosphat existiert in drei möglichen Formen (der Resonanz) und das macht es recht stabil. Darüber hinaus überlebt das Hydroxyethyl-Thiamin-Pyrophosphat in anionischer Form lange genug, um mit der Disulfidbrücke des Lipoamids (zweiter katalytischer Cofaktor der Pyruvat-Dehydrogenase) wechselwirken zu können; die Disulfidbrücke ist ein schwingender Arm (sie befindet sich am Ende einer langen flexiblen Kette) und können sich im Enzymkomplex von einem katalytischen Zentrum zum anderen bewegen.
Dann bindet das Lipoamid über die Disulfidbrücke das Hydroxyethyl-Thiamin-Pyrophosphat: Acetyllipoamid wird erhalten.Der gerade beschriebene ist der erste Schritt einer Transacetylierungsreaktion, die durch das erste Enzym des Pyruvat-Dehydrogenase-Komplexes katalysiert wird; in dieser Phase wurde eine Bindung zwischen der Hydroxylgruppe und dem Thiaminpyrophosphat gebrochen, das in seine ursprüngliche Form zurückkehrte: Es fand eine Redoxreaktion statt, bei der die Disulfidbrücke als Oxidationsmittel (die beiden Schwefelatome reduziert) in den Vergleichen in der Hydroxylgruppe wirkte, die zu Acetyl oxidiert.
Nach dieser Phase bewegt sich der schwingende Arm des Lipoamids und nähert sich dem zweiten Enzym der Pyruvat-Dehydrogenase, das die eigentliche Transacetylase-Aktivität ausführt, indem es die Acetylgruppe mitnimmt: die zweite Phase der durch das zweite Enzym katalysierten Transacetylierungsreaktion findet statt; Auf diese Weise haben wir das Acetyl-Coenzym A erhalten FAD / FADH2 kann als Redoxpaar in zwei verschiedenen monoelektronischen Stufen oder in einer einzelnen bielektronischen Stufe fungieren.
Das FADH2 gibt seine Elektronen sofort an das NAD + ab, wodurch FAD und NADH + H + erhalten werden.
Acetyl-Coenzym A, wie beschrieben erhalten, ist das Ausgangsprodukt für den Krebs-Zyklus (oder Zyklus der Tricarbonsäuren).